O Mesozóico é uma era do tempo geológico compreendida entre 252 – 66 milhões de anos (m.a.) que começa após a maior extinção em massa da história e se divide em três períodos, de acordo com aspectos paleobiogeográficos, sendo esses: Triássico (252 – 201 m.a.), Jurássico (201 – 143 m.a.) e o Cretáceo (143 – 66 m.a.) (Cohen et al., 2013), tendo fim com a queda de um meteorito que se chocou com a Terra onde hoje é a América do Norte (Schulte et al., 2010), culminando na extinção de diversos grupos de plantas e animais.
Ainda no começo dessa era existia a Pangéia, um supercontinente formado por todas as massas de terras então emersas que durou desde o Carbonífero Superior até o Triássico Médio, sendo esse, banhado de todos os lados pelo oceano Pantalassa (Wopfner & Jin, 2009). 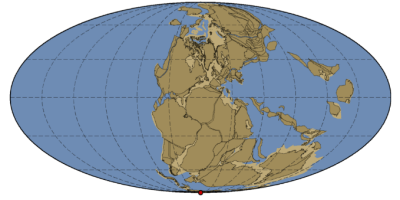 Devido a essa aglomeração de terras, o Mesozóico é marcado por uma relativa estabilidade climática, com o Triássico e o Jurássico sendo bastante áridos, sobretudo no interior, devido ao efeito de continentalidade (Muñoz et al., 1985), enquanto que no Cretáceo Inferior houve um período mais úmido, sobretudo perto do Mar de Tethys (Hallam, 1985).
Devido a essa aglomeração de terras, o Mesozóico é marcado por uma relativa estabilidade climática, com o Triássico e o Jurássico sendo bastante áridos, sobretudo no interior, devido ao efeito de continentalidade (Muñoz et al., 1985), enquanto que no Cretáceo Inferior houve um período mais úmido, sobretudo perto do Mar de Tethys (Hallam, 1985).
Fig. 1: Mapa da Pangéia durante o Triássico. Créditos: Eikeskog1225.
Embora seja conhecido popularmente como a ‘’Era dos dinossauros’’, inúmeros outros grupos de organismos são característicos do Mesozóico, desde vertebrados terrestres como pterossauros (Baron, 2021), rincossauros (Ezcurra et al., 2016) e os primeiros mamíferos (Kermack & Kermack, 1984), diversas linhagens de répteis marinhos como os ictiossauros, plesiossauros e mosassauros (Motani, 2009), invertebrados como os amonites (Kennedy, 1997) e as primeiras plantas angiospermas (Axelrod, 1970).

Fig. 2: Paleoambiente terrestre do Mesozóico mostrando dinossauros e mamíferos convivendo. Créditos: Mark Witton, Steven C. Sweetman, Grant Smith e David M. Martill.

Fig. 3: Paleoambiente marinho do Mesozóico mostrando interações entre mosassauros, amonites e peixes ósseos. Créditos: Andrey Atuchin.
Triássico
O Triássico é o primeiro período da era Mesozóica, que começa após a extinção em massa mais devastadora do éon Fanerozóico, a extinção do Permo-Triássico, também conhecida como ‘’A grande morte’’, que dizimou cerca de 90% das espécies marinhas e 70% das terrestres (Erwin, 1994 e 1995), causada pela acidificação, anóxia e elevação na temperatura dos oceanos (Wignall & Bond, 2024) e um intenso vulcanismo responsável por um efeito estufa e aquecimento global catastróficos (Grard et al., 2005).
Durante esse período, a Pangéia já apresentava alguns pontos de rifteamento que posteriormente contribuíram para a separação completa da mesma (Frizon de Lamotte, 2015; Golonka, 2007), dando origem a dois supercontinentes: Laurásia (ao norte), incluindo terras que hoje são a América do Norte e Eurásia, e Gondwana (ao sul), que inclui o que atualmente é a América do Sul, África, Oriente Médio, Subcontinente Indiano, Austrália e Antártica (Gargaud et al., 2015).
Em geral, o Triássico foi um período bastante quente e seco, embora tenham existido episódios mais úmidos, como foi o Evento Pluvial do Carniano (Benton et al., 2018), que aconteceu há cerca de 232 m.a. e proporcionou uma irradiação evolutiva de vários grupos de vertebrados que depois se tornaram dominantes em ambientes terrestres durante o resto da era Mesozóico (Dal Corso et al., 2020).

Fig. 4: Paleoambiente do Triássico mostrando rincossauros (à frente) e Prestosuchus (ao fundo). Créditos: Gabriel Ugueto.
A paleoflora do Triássico é formada, principalmente, pela assembléia denominada Flora de Dicroidium, que consiste em plantas adaptadas a regiões mais áridas, condizentes com as adaptações que apresentam, como cutícula mais grossa e anéis de crescimento bem marcados (Colombi et al., 2011; Sommer et al., 1999). Entre os principais grupos de plantas dessa assembléia estão as Pteridospermales (Dicroidium), Equisetales (cavalinhas), Cycadales (cicas), Ginkgoales (atualmente representada apenas pela espécie Ginkgo biloba) e as Pinales (coníferas), entre outros (Chatterjee et al., 2013).

Fig. 5: Paleoflora de Dicroidium presente no Triássico. Créditos: Michael Rothman.
Formação Santa Maria
A Formação Santa Maria, datada do período Carniano (237 – 227 m.a.) (Zerfass et al., 2005), está localizado na Depressão Central do estado do Rio Grande do Sul, entre os municípios de Mata e Venâncio Aires e apresenta uma grande variedade de fósseis de vertebrados (Abdala & Dornelles, 2002), entre os quais se destacam os primeiros dinossauros conhecidos (Langer et al., 2010; Marsola et al., 2018).
Essa formação está dividida em quatro biozonas (intervalos estratigráficos marcados pela ocorrência de um taxon ou linhagem específicos (MacLeod, 2005)), sendo elas, da mais antiga à mais recente: Zona de Associação (ZA) de Dinodontosaurus (com abundância de fósseis de cinodontes como o Massetognathus e arcossauros como o Prestosuchus), ZA de Santacruzodon (composta por cinodontes não-mamaliformes (Horn et al., 2014), presentes também na ZA anterior), ZA de Hyperodapedon (abrangendo inúmeras linhagens, com destaque para o rincossauro Hyperodapedon, Lagerpetidae como Ixalerpeton e o recém descoberto Venetoraptor (Müller et al., 2023), além de dinossauros como Buriolestes e Bagualosaurus) e ZA de Riograndia (rico em cinodontes como Brasilodon, Procolophonidae como o Soturnia e dinossauros como Sacisaurus e Guaibasaurus) (Barberena, 1977; Schultz et al., 2020).

Fig. 6: Prestosuchus (devorando dicinodonte) e um grupo de Parvosuchus (à frente).
Créditos: Matheus Fernandes.
Primeiros dinossauros
Os fósseis mais antigos conhecidos, que com certeza pertencem à dinossauros, são encontrados na Formação Santa Maria (Langer et al., 2010; Marsola et al., 2018), seguidos, provavelmente, pela Formação Ischigualasto, na Argentina (Martínez et al., 2012).
Os dinossauros formam um grupo de Archosauria (Benton & Clark, 1988) e são reconhecidos, entre outras características, por uma pélvis com um acetábulo aberto para o encaixe do fêmur, por ossos do tornozelo (astrágalo e calcâneo) mais reforçados (Blanco et al., 2020; Brusatte, 2012), além da presença de protopenas já nas primeiras linhagens (Lingham-Soliar, 2010; Xu & Barrett, 2025).
São divididos em dois grupos segundo a direção em que aponta o osso do púbis (Macaluso & Tschopp, 2018): Ornithischia, com um púbis apontando para trás e um bico córneo formado a partir do osso pré-dentário (ex: Triceratops, Parasaurolophus, Stegosaurus e Ankylosaurus) (Ferigolo & Langer, 2007; Norman et al., 2004); e Saurischia, com o púbis apontando para a frente; sendo esse segundo grupo subdividido em Sauropodomorpha, quadrúpedes preferencialmente herbívoros, inicialmente de pequeno porte (ex: Plateosaurus) (Yates, 2012), mas que ao longo do Mesozóico darão origem aos maiores animais terrestres da história (ex: Brachiosaurus e Diplodocus) (Sander et al., 2011); Theropoda, formado por carnívoros bípedes (ex: Allosaurus, Tyrannosaurus, Velociraptor e Archaeopteryx) (Yates, 2005) e que, posteriormente, darão origem ao único grupo de dinossauros vivos: as Aves (Abourachid & Höfling, 2012; Clarke & Middleton, 2006); além dos Herrerasauridae, uma linhagem de dinossauros bípedes e carnívoros, convergentemente similares aos Theropoda, porém restritos ao Triássico da América do Sul (ex: Gnathovorax e Staurikosaurus) (Benedetto, 1973; Garcia et al., 2023).
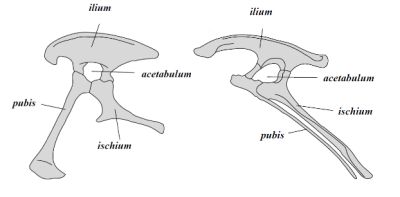
Fig. 7: Diferenças na bacia entre Saurischia (esq.) e Ornithischia (dir.). Créditos: AdmiralHood.
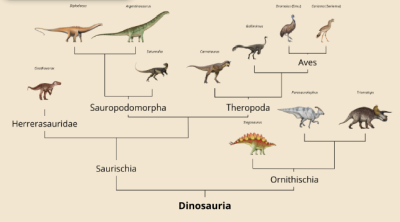
Fig. 8: Árvore filogenética baseada nos estudos de Langer et al., 2017. Créditos: Vários artistas (Apêndice 1).
Referências
Abdala, F., Dornelles, J. E. F. 2002. Herpetofauna Permotriássica no Rio Grande do Sul. p.111-120. in:Os Répteis do Rio Grande do Sul. In: LEMA, Thales de (Org). Os Répteis do Rio Grande do Sul.2002, 166p. EdiPUCRS.Porto Alegre.
Abourachid, A., & Höfling, E. (2012). The legs: a key to bird evolutionary success. Journal of Ornithology, 153(Suppl 1), 193-198.
Axelrod, D. I. (1970). Mesozoic paleogeography and early angiosperm history. The Botanical Review, 36(3), 277-319.
Barberena, M. C. (1977). Bioestratigrafia preliminar da formação Santa Maria. Pesquisas em Geociências, 7(7), 111-129.
Baron, M. G. (2021). The origin of Pterosaurs. Earth-Science Reviews, 221, 103777.
Benedetto, J. L. (1973). Herrerasauridae, nueva familia de saurisquios triasicos. Ameghiniana, 10(1), 89-102.
Benton, M. J., Bernardi, M., & Kinsella, C. (2018). The Carnian Pluvial Episode and the origin of dinosaurs. Journal of the Geological Society, 175(6), 1019-1026.
Benton, M. J., & Clark, J. M. (1988). Archosaur phylogeny and the relationships of the Crocodylia. The phylogeny and classification of the tetrapods, 1, 295-338.
Blanco, M.V.F., Ezcurra, M.D. & Bona, P. New embryological and palaeontological evidence sheds light on the evolution of the archosauromorph ankle. Sci Rep 10, 5150 (2020).
Brusatte, S. L. (2012). Dinosaur paleobiology. John Wiley & Sons.
Chatterjee, S., Tewari, R., & Agnihotri, D. (2013). A Dicroidium flora from the Triassic of Allan Hills, South Victoria Land, Transantarctic Mountains, Antarctica. Alcheringa: An Australasian Journal of Palaeontology, 37(2), 209-221.
Clarke, J., & Middleton, K. (2006). Bird evolution. Current Biology, 16(10), R350-R354.
Cohen, K.M., Finney, S.C., Gibbard, P.L. & Fan, J.-X. (2013; updated) The ICS International Chronostratigraphic Chart. Episodes 36: 199-204.
Colombi, C. E., Montañez, I. P., & Parrish, J. T. (2011). Registro de la Relación Isotópica de Carbono en la paleoflora de la Formación Ischigualasto (Triásico Superior), Noroeste Argentina: Implicancias paleoatmosféricas.
Dal Corso, J., Bernardi, M., Sun, Y., Song, H., Seyfullah, L. J., Preto, N., … & Benton, M. J. (2020). Extinction and dawn of the modern world in the Carnian (Late Triassic). Science Advances, 6(38), eaba0099.
Erwin, D. H. (1994). The permo–triassic extinction. Nature, 367(6460), 231-236.
Erwin, D. H. (1995). The great Paleozoic crisis: life and death in the Permian. (No Title).
Ezcurra, M. D., Montefeltro, F., & Butler, R. J. (2016). The early evolution of rhynchosaurs. Frontiers in Ecology and Evolution, 3, 142.
Ferigolo, J., & Langer, M. C. (2007). A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone. Historical Biology, 19(1), 23-33.
Frizon de Lamotte, D., Fourdan, B., Leleu, S., Leparmentier, F., & de Clarens, P. (2015). Style of rifting and the stages of Pangea breakup. Tectonics, 34(5), 1009-1029.
Garcia, M. S., Müller, R. T., Ghilardi, A. M., & Paes Neto, V. (2023). Gnathovorax cabreirai, um dinossauro excepcionalmente bem preservado do Triássico superior do sul do Brasil.
Gargaud, M., Irvine, W. M., Amils, R., Claeys, P., Cleaves, H. J., Gerin, M., … & Viso, M. (Eds.). (2015). Encyclopedia of astrobiology. Berlin, Heidelberg: Springer Berlin Heidelberg.
Golonka, J. (2007). Late Triassic and Early Jurassic palaeogeography of the world. Palaeogeography, Palaeoclimatology, Palaeoecology, 244, 297-307.
Grard, A., Francois, L. M., Dessert, C., Dupré, B., & Godderis, Y. (2005). Basaltic volcanism and mass extinction at the Permo-Triassic boundary: environmental impact and modeling of the global carbon cycle. Earth and Planetary Science Letters, 234(1-2), 207-221.
Hallam, A. (1985). A review of Mesozoic climates. Journal of the Geological Society, 142(3), 433-445.
Horn, B. L. D., Melo, T. M., Schultz, C. L., Philipp, R. P., Kloss, H. P., & Goldberg, K. (2014). A new third-order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. Journal of South American Earth Sciences, 55, 123-132.
Kennedy, W. J. (1977). Ammonite evolution. In Developments in palaeontology and stratigraphy (Vol. 5, pp. 251-304). Elsevier. Kermack, D. M., & Kermack, K. A. (1984). The first mammals. In The Evolution of Mammalian Characters (pp. 48-65). Boston, MA: Springer US.
Langer, M. C., Ezcurra, M. D., Bittencourt, J. S., & Novas, F. E. (2010). The origin and early evolution of dinosaurs. Biological Reviews, 85(1), 55-110.
Langer, M. C., Ezcurra, M. D., Rauhut, O. W., Benton, M. J., Knoll, F., McPhee, B. W., … & Brusatte, S. L. (2017). Untangling the dinosaur family tree. Nature, 551(7678), E1-E3.
Lingham-Soliar, T. Dinosaur protofeathers: pushing back the origin of feathers into the Middle Triassic?. J Ornithol 151, 193–200 (2010).
Macaluso, L., & Tschopp, E. (2018). Evolutionary changes in pubic orientation in dinosaurs are more strongly correlated with the ventilation system than with herbivory. Palaeontology, 61(5), 703-719.
MacLeod, N. (2005). Principles of stratigraphy. Encyclopedia of Geology, 2005, 295-307.
Marsola, J. C., Bittencourt, J. S., Butler, R. J., Da Rosa, Á. A., Sayão, J. M., & Langer, M. C. (2018). A new dinosaur with theropod affinities from the Late Triassic Santa Maria Formation, south Brazil. Journal of Vertebrate Paleontology, 38(5).
Martínez, R. N., Apaldetti, C., Alcober, O. A., Colombi, C. E., Sereno, P. C., Fernandez, E., … & Abelin, D. (2012). Vertebrate succession in the Ischigualasto Formation. Journal of Vertebrate paleontology, 32(sup1), 10-30.
Motani, R. (2009). The evolution of marine reptiles. Evolution: Education and Outreach, 2(2), 224-235.
Müller, R.T., Ezcurra, M.D., Garcia, M.S. et al. New reptile shows dinosaurs and pterosaurs evolved among diverse precursors. Nature 620, 589–594 (2023).
Muñoz, J. M., Alvarado, J. M. G., & Aldeanueva, R. A. (1985). Características de la continentalidad en el clima de la España peninsular. Geographica, 27, 41.
Norman, D. B., Witmer, L. M., & Weishampel, D. B. (2004). Basal ornithischia. The dinosauria, 2, 325-334.
Sander, P. M., Christian, A., Clauss, M., Fechner, R., Gee, C. T., Griebeler, E. M., … & Witzel, U. (2011). Biology of the sauropod dinosaurs: the evolution of gigantism. Biological Reviews, 86(1), 117-155.
Schulte, P., Alegret, L., Arenillas, I., Arz, J. A., Barton, P. J., Bown, P. R., … & Willumsen, P. S. (2010). The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary. Science, 327(5970), 1214-1218.
Schultz, C. L., Martinelli, A. G., Soares, M. B., Pinheiro, F. L., Kerber, L., Horn, B. L., … & Melo, T. P. (2020). Triassic faunal successions of the Paraná Basin, southern Brazil. Journal of South American Earth Sciences, 104, 102846.
Sommer, M. G., Klepzig , M. C., Iannuzzi, R., & Alves, L. S. (1999). A Flora Dicroidium no Rio grande do Sul: implicações bioestratigráficas. Pesquisas em Geociências, 26(1), 3-9.
Wignall, P. B., & Bond, D. P. (2024). The great catastrophe: causes of the Permo-Triassic marine mass extinction. National Science Review, 11(1), nwad273.
Wopfner, H., & Jin, X. C. (2009). Pangea Megasequences of Tethyan Gondwana-margin reflect global changes of climate and tectonism in Late Palaeozoic and Early Triassic times—A review. Palaeoworld, 18(2-3), 169-192.
Xu, X., & Barrett, P. M. (2025). The origin and early evolution of feathers: implications, uncertainties and future prospects. Biology Letters, 21(2), 20240517.
Yates, A. M. (2005). A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods. Palaeontologia africana, 41, 105-122.
Yates, A. M. (2012). Basal Sauropodomorpha: the “Prosauropods.”. The Complete Dinosaur,, 425-443.
Zerfass, H., Chemale Jr, F., & Lavina, E. (2005). Tectonic control of the Triassic Santa Maria Supersequence of the Paraná Basin, southernmost Brazil, and its correlation to the Waterberg Basin, Namibia. Gondwana Research, 8(2), 163-176.
Apêndice 1:
| Cariama (Seriema) | Nicolas Huet / Jean Gabriel Prêtre |
| Dromaius (Emu) | ARC CoE CABAH |
| Gallimimus | PaleoNeolitic |
| Carnotaurus | Fred Wierum |
| Argentinosaurus | TotalDino |
| Diplodocus | Fred Wierum |
| Saturnalia | Caetano Soares |
| Gnathovorax | Caetano Soares |
| Triceratops | Camus Altamirano |
| Parasaurolophus | Debivort |
| Stegosaurus | TotalDino |
| Composição | Daniel Dias |